Type I collagen synthesis in the context of osteogenesis imperfecta (WP4786)
Homo sapiens

{kind=link}
{kind=link}
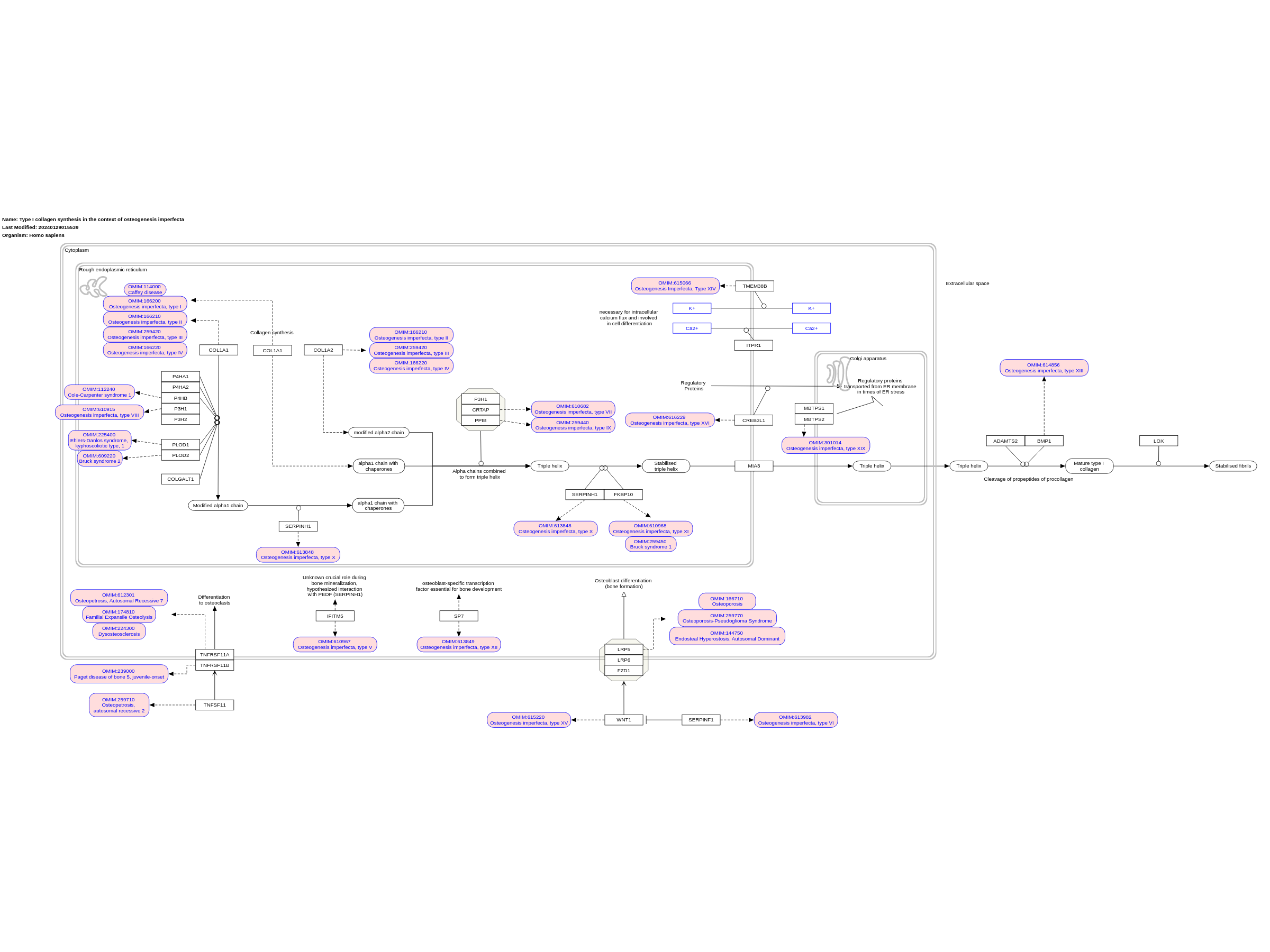
Type I collagen — the major protein component of the extracellular matrix in bone, skin and tendon — is mainly secreted by osteoblasts, dermal fibroblasts and tenocytes. Despite the relatively simple structure of the collagen triple helix, the biosynthesis of type I procollagen is extremely complex, involving multiple steps and requiring an ensemble of proteins for post-translational modifications, folding, transport, secretion and quality control. The COL1A1 and COL1A2 transcripts are translated in the rough endoplasmic reticulum (rER), and the α(I)‑chains undergo a series of post-translational modifications. Helical prolines in position Y of the Gly-Xaa-Yaa repeat are hydroxylated in position C4 by prolyl 4‑hydroxylase 1 (P4H1), whereas specific prolines in the X positions are 3‑hydroxylated by P3H1 and P3H2. Some lysine residues are hydroxylated by lysyl hydroxylases (LH1 and LH2, encoded by PLOD1 and PLOD2 respectively), and glycosylation of hydroxylysines into galactosyl-hydroxylysine and glucosyl-galactosyl-hydroxylysine is catalysed by procollagen galactosyltransferase 1 and procollagen glucosyltransferase 1. After synthesis of the carboxy-terminal propeptide, it forms intra-chain disulfide bonds and remains attached to the rER membrane. Selection and association of the correct chains into a triple helix occur by diffusion of the C-propeptides attached to the rER membrane. A nucleus for triple helix formation is formed that staggers the chains in the correct order and initiates triple helix formation. Protein disulfide isomerase catalyses inter-chain disulfide bond formation, which stabilize the folding nucleus. Hydroxylation of proline residues and some lysine residues continues and triple helix formation proceeds from the C-terminal end towards the amino-terminal end. During this phase, 65 kDa FK506‑binding protein (FKBP65; encoded by FKBP10) and a complex formed by P3H1 — CRTAP– PPIase B (peptidyl-prolyl cis–trans isomerase B) — seem to play a crucial part. The complex is involved in the Hydroxylation of proline 986 of the collagen α1(I)-chain and α1(II)-chain and proline 707 of the α2(I)-chain, which are thought to be important for supramolecular assembly of collagen fibrils and to serve as binding sites for chaperones or small leucine-rich proteoglycans. Beyond its prolyl 3‑hydroxylase activity, the complex functions as a PPIase and chaperone for collagen folding. Indeed, the fast propagation of the triple helix requires the isomerization of cis peptide bonds that convert proline residues into trans configuration, mainly by PPIase B24. When most of the helix is folded, the N-propeptides associate and form the small triple helix within this domain. The newly formed triple helix is stabilized by serpin H1 (also known as HSP47; encoded by SERPINH1) and FKBP65. Further modifications occur during transport from the rER to the Golgi apparatus in special coat protein complex vesicles that contain melanoma inhibitory activity protein 3 (also known as TANGO1, encoded by MIA) and through the Golgi stack by cisternal maturation. Serpin H1 also has binding sites along the helical portion of the molecule and assists shuttling of folded collagen into the cis-Golgi. These biosynthetic steps depend on a proper rER environment (for example, optimal calcium levels and redox potential), and the quality-control mechanisms can lead to the activation of the unfolded protein response using the ER-associated degradation pathway or the autophagy-mediated lysosomal degradation system to eliminate molecules that were not properly folded. Once secreted, the propeptides of procollagen are cleaved by a disintegrin and metalloproteinase with thrombospondin motifs 2 (ADAMTS2) and bone morphogenetic protein 1 (BMP1) into mature type I collagen. This initiates collagen fibre formation and these fibrils are stabilized by crosslink formation, in which certain lysine and hydroxylysine residues in the triple helix and the telopeptides are oxidized by lysyl oxidases and converted into allysine and hydroxyallysine. These residues then initially form divalent crosslinks that convert into mature trivalent pyridinoline and pyrrole crosslinks to stabilize the fibril structure in tissues. Bone formation consists of the secretion of bone extracellular matrix components (mainly type I collagen) by osteoblasts. The unmineralized bone matrix (osteoid) subsequently becomes mineralized. In addition, osteoblasts and osteocytes release many cytokines, including receptor activator of nuclear factor-κB ligand (RANKL; also known as TNFSF11) and osteoprotegerin (OPG, encoded by TNFRSF11B), which regulate bone resorption by osteoclasts. RANKL acts on osteoclast precursor cells by binding to receptor activator of nuclear factor-κB (RANK; also known as TNFRSF11A) on their surface, thereby favouring their differentiation to osteoclasts. OPG, by interacting with RANKL, prevents the binding of RANKL to RANK. Some osteoblasts become embedded in the mineralized bone matrix and differentiate to osteocytes, which produce, among other factors, sclerostin, an inhibitor of the WNT pathway that is known to stimulate bone formation by stimulating osteoblast activity. Linked with a dotted arrow to the GeneProduct nodes are diseases caused by mutation in the respective gene. Adapted from [5]
For a description of pathway objects, see the WikiPathways Legend.
Authors
Ritchie Lee , Kristina Hanspers , Egon Willighagen , Andreas Zankl , Eric Weitz , and Martina Summer-KutmonActivity

Discuss this pathway
Check for ongoing discussions or start your own.
Cited In
- Blood biomarkers representing maternal-fetal interface tissues used to predict early-and late-onset preeclampsia but not COVID-19 infection (2022).
- Transcriptome analysis of newly established carboplatin-resistant ovarian cancer cell model reveals genes shared by drug resistance and drug-induced EMT (2023).
Are you planning to include this pathway in your next publication? See How to Cite and add a link here to your paper once it's online.
Organisms
Homo sapiensCommunities
Skeletal DysplasiaAnnotations
Disease Ontology
osteogenesis imperfectaPathway Ontology
disease pathwayReferences
- Prolyl 3-hydroxylase 1, enzyme characterization and identification of a novel family of enzymes. Vranka JA, Sakai LY, Bächinger HP. J Biol Chem. 2004 May 28;279(22):23615–21. PubMed Europe PMC Scholia
- CRTAP is required for prolyl 3- hydroxylation and mutations cause recessive osteogenesis imperfecta. Morello R, Bertin TK, Chen Y, Hicks J, Tonachini L, Monticone M, et al. Cell. 2006 Oct 20;127(2):291–304. PubMed Europe PMC Scholia
- Location of 3-hydroxyproline residues in collagen types I, II, III, and V/XI implies a role in fibril supramolecular assembly. Weis MA, Hudson DM, Kim L, Scott M, Wu JJ, Eyre DR. J Biol Chem. 2010 Jan 22;285(4):2580–90. PubMed Europe PMC Scholia
- New perspectives on osteogenesis imperfecta. Forlino A, Cabral WA, Barnes AM, Marini JC. Nat Rev Endocrinol. 2011 Jun 14;7(9):540–57. PubMed Europe PMC Scholia
- Osteogenesis imperfecta. Marini JC, Forlino A, Bächinger HP, Bishop NJ, Byers PH, Paepe AD, et al. Nat Rev Dis Primers. 2017 Aug 18;3:17052. PubMed Europe PMC Scholia