Nutrient control of ribosomal gene expression (WP2869)
Saccharomyces cerevisiae

{kind=link}
{kind=link}
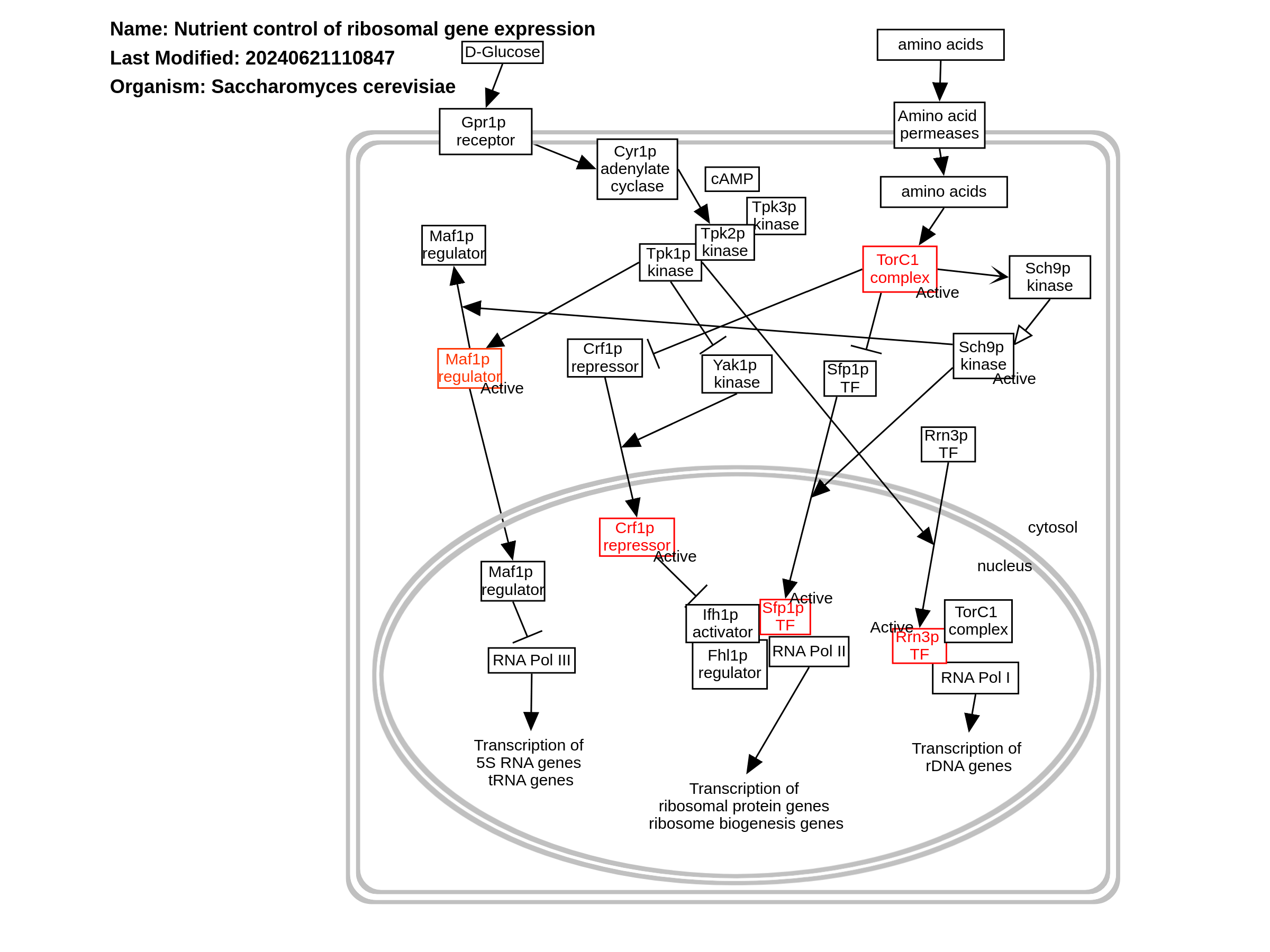
Ribosomes are highly conserved large ribonucleoprotein (RNP) particles, consisting in yeast of a large 60S subunit and a small 40S subunit, that perform protein synthesis. Yeast ribosomes contain one copy each of four ribosomal RNAs (5S, 5.8S, 18S, and 25S; produced in two separate transcripts encoded within the rDNA repeat present as hundreds of copies on Chromosome 12) and 79 different ribosomal proteins (r-proteins), which are encoded by 137 different genes scattered about the genome, 59 of which are duplicated. The 60S subunit contains 46 proteins and three RNA molecules: 25S RNA of 3392 nt, hydrogen bonded to the 5.8S RNA of 158 nt and associated with the 5S RNA of 121 nt. The 40S subunit has a single 18S RNA of 1798 nt and 33 proteins. All yeast ribosomal proteins have a mammalian homolog. PMID: 10690410, PMID: 22884264, PMID: 9421530, PMID: 9396790 In a rapidly growing yeast cell, 60% of total transcription is devoted to ribosomal RNA, and 50% of RNA polymerase II transcription and 90% of mRNA splicing are devoted to the production of mRNAs for r-proteins. Coordinate regulation of the rRNA genes and 137 r-protein genes is affected by nutritional cues and a number of signal transduction pathways that can abruptly induce or silence the ribosomal genes, whose transcripts have naturally short lifetimes, leading to major implications for the expression of other genes as well. The expression of some r-protein genes is influenced by Abf1p, and most are directly induced by binding of Rap1p to their promoters, which excludes nucleosomes and recruits Fhl1p and Ifh1p to drive transcription. PMID: 10409730, PMID: 12509467, PMID: 10542411, PMID: 2207166, PMID: 16782874 Ribosome synthesis is under nutrient control. Synthesis of the four rRNAs and 79 different proteins in equimolar amounts is one of the most energetically expensive cellular processes, and must be coordinated together. Ribosome biogenesis requires all three RNA polymerases: Pol I for rRNA genes, Pol II for ribosomal protein genes, and Pol III for tRNA and 5S RNA genes. Therefore, sensing quality and quantity of available nutrients is key in the regulation of ribosome biogenesis. PMID: 15489289, PMID: 18303986 Formation of transcription initiation complexes at the rDNA promoter depends on the association of RNA Pol I with the Rrn3p transcription factor, which is regulated by phosphorylation/dephosphorylation of Rrn3p. During normal conditions, a surface serine patch on Rrn3p is not phosphorylated, enabling it to bind RNA Pol I for efficient rDNA transcription. In stress conditions, the surface serine patch undergoes phosphorylation, which impairs the interaction of Rrn3p with RNA Pol I, repressing Pol I transcription, and thereby also reducing ribosome production and cell growth. The presence of glucose results in increased expression of RRN3. In cells treated with rapamycin, Rrn3p is subject to proteasome-dependent degradation, reducing the cellular amount of transcription-initiation competent RNA Pol I - Rrn3p complexes. PMID: 19796927, PMID: 8670901, PMID: 20421203, PMID: 20154141, PMID: 18084032, PMID: 11717393, PMID: 14595104 Ribosomal protein genes and ribosomal biogenesis genes are transcribed by RNA polymerase II. The activation of these promoters requires the forkhead-like transcription factor, Fhl1p, the acitvity of which is subject to nutrient regulation via the Ifh1 activator and the Crf1 repressor. Under lack of stress, the TorC1 complex retains Crf1p in the cytoplasm in a dephosphorylated, inactive state. Upon nutrient deprivation, the PKA-responsive Yak1 kinase phosphorylates Crf1p, resulting in its translocation to the nucleus, where Crf1p competes with the Ifh1 activator to bind Fhl1p. The transcription factor Sfp1p also mediates TorC1 regulation of ribosomal protein and ribosomal biogenesis genes. Interaction of Sfp1p with TorC1 reduces Sch9p phosphorylation, resulting in a negative feedback loop. PMID: 15620355, PMID: 19796927 Transcription by RNA polymerase III is negatively regulated by Maf1, a highly conserved repressor. Both the localization and activity of Maf1p are regulated by phosphorylation at various sites, mediated by TORC1, protein kinase A (PKA), and Sch9p. In its dephosphorylated state, Maf1p binds the N-terminus of the Rpc160p subunit of Pol III to prevent closed-complex formation. Maf1p is maintained in the cytoplasm during vegetative growth via PKA- or Sch9p-mediated phosphorylation. Dephosphorylation allows translocation of Maf1p to the nucleus and nucleolus under stress conditions, which shuts down RNA Pol III transcription.Phosphorylation of Maf1p by CK2 occurs at promoters, which releases Maf1p from chromatin, liberating RNA Pol III from inhibition. PMID: 19684113, PMID: 19299514, PMID: 21383183, PMID: 17005718, PMID: 22810236
For a description of pathway objects, see the WikiPathways Legend.
Authors
Stacia , Bart Smeets , Martina Summer-Kutmon , and Egon WillighagenActivity

Discuss this pathway
Check for ongoing discussions or start your own.
Cited In
Are you planning to include this pathway in your next publication? See How to Cite and add a link here to your paper once it's online.
Organisms
Saccharomyces cerevisiaeCommunities
Annotations
Pathway Ontology
regulatory pathway signaling pathway| Label | Type | Compact URI | Comment |
|---|---|---|---|
| D-Glucose | Metabolite | hmdb:HMDB0000122 | |
| amino acids | Metabolite | chebi:33709 | |
| cAMP | Metabolite | hmdb:HMDB0000058 | |
| Rrn3pTF | Protein | sgd:S000001608 | |
| Sfp1pTF | Protein | sgd:S000004395 | |
| Amino acidpermeases | Protein | wikidata:Q4746432 | |
| Maf1pregulator | Protein | sgd:S000002412 | |
| Sch9pkinase | Protein | sgd:S000001248 | |
| Gpr1preceptor | Protein | sgd:S000002193 | |
| Tpk1pkinase | Protein | sgd:S000003700 | |
| Tpk3pkinase | Protein | sgd:S000001649 | |
| Yak1pkinase | Protein | sgd:S000003677 | |
| Crf1prepressor | Protein | sgd:S000002631 | |
| Fhl1pregulator | Protein | sgd:S000006308 | |
| Ifh1pactivator | Protein | sgd:S000004213 | |
| Tpk2pkinase | Protein | sgd:S000006124 | |
| Cyr1padenylatecyclase | Protein | sgd:S000003542 |
References
- Multifunctional DNA-binding proteins mediate concerted transcription activation of yeast ribosomal protein genes. Mager WH, Planta RJ. Biochim Biophys Acta. 1990 Aug 27;1050(1–3):351–5. PubMed Europe PMC Scholia
- RRN3 gene of Saccharomyces cerevisiae encodes an essential RNA polymerase I transcription factor which interacts with the polymerase independently of DNA template. Yamamoto RT, Nogi Y, Dodd JA, Nomura M. EMBO J. 1996 Aug 1;15(15):3964–73. PubMed Europe PMC Scholia
- A new nomenclature for the cytoplasmic ribosomal proteins of Saccharomyces cerevisiae. Mager WH, Planta RJ, Ballesta JG, Lee JC, Mizuta K, Suzuki K, et al. Nucleic Acids Res. 1997 Dec 15;25(24):4872–5. PubMed Europe PMC Scholia
- Three-dimensional structure of the yeast ribosome. Verschoor A, Warner JR, Srivastava S, Grassucci RA, Frank J. Nucleic Acids Res. 1998 Jan 15;26(2):655–61. PubMed Europe PMC Scholia
- Transcriptional elements involved in the repression of ribosomal protein synthesis. Li B, Nierras CR, Warner JR. Mol Cell Biol. 1999 Aug;19(8):5393–404. PubMed Europe PMC Scholia
- The economics of ribosome biosynthesis in yeast. Warner JR. Trends Biochem Sci. 1999 Nov;24(11):437–40. PubMed Europe PMC Scholia
- Ribosome synthesis in Saccharomyces cerevisiae. Venema J, Tollervey D. Annu Rev Genet. 1999;33:261–311. PubMed Europe PMC Scholia
- Differential roles of phosphorylation in the formation of transcriptional active RNA polymerase I. Fath S, Milkereit P, Peyroche G, Riva M, Carles C, Tschochner H. Proc Natl Acad Sci U S A. 2001 Dec 4;98(25):14334–9. PubMed Europe PMC Scholia
- Autoregulation in the biosynthesis of ribosomes. Zhao Y, Sohn JH, Warner JR. Mol Cell Biol. 2003 Jan;23(2):699–707. PubMed Europe PMC Scholia
- Tor pathway regulates Rrn3p-dependent recruitment of yeast RNA polymerase I to the promoter but does not participate in alteration of the number of active genes. Claypool JA, French SL, Johzuka K, Eliason K, Vu L, Dodd JA, et al. Mol Biol Cell. 2004 Feb;15(2):946–56. PubMed Europe PMC Scholia
- What better measure than ribosome synthesis? Rudra D, Warner JR. Genes Dev. 2004 Oct 15;18(20):2431–6. PubMed Europe PMC Scholia
- Fine-structure analysis of ribosomal protein gene transcription. Zhao Y, McIntosh KB, Rudra D, Schawalder S, Shore D, Warner JR. Mol Cell Biol. 2006 Jul;26(13):4853–62. PubMed Europe PMC Scholia
- Protein kinase A regulates RNA polymerase III transcription through the nuclear localization of Maf1. Moir RD, Lee J, Haeusler RA, Desai N, Engelke DR, Willis IM. Proc Natl Acad Sci U S A. 2006 Oct 10;103(41):15044–9. PubMed Europe PMC Scholia
- Site specific phosphorylation of yeast RNA polymerase I. Gerber J, Reiter A, Steinbauer R, Jakob S, Kuhn CD, Cramer P, et al. Nucleic Acids Res. 2008 Feb;36(3):793–802. PubMed Europe PMC Scholia
- How Saccharomyces responds to nutrients. Zaman S, Lippman SI, Zhao X, Broach JR. Annu Rev Genet. 2008;42:27–81. PubMed Europe PMC Scholia
- Regulation of RNA polymerase III transcription involves SCH9-dependent and SCH9-independent branches of the target of rapamycin (TOR) pathway. Lee J, Moir RD, Willis IM. J Biol Chem. 2009 May 8;284(19):12604–8. PubMed Europe PMC Scholia
- Characterization of the rapamycin-sensitive phosphoproteome reveals that Sch9 is a central coordinator of protein synthesis. Huber A, Bodenmiller B, Uotila A, Stahl M, Wanka S, Gerrits B, et al. Genes Dev. 2009 Aug 15;23(16):1929–43. PubMed Europe PMC Scholia
- Growth control and ribosome biogenesis. Lempiäinen H, Shore D. Curr Opin Cell Biol. 2009 Dec;21(6):855–63. PubMed Europe PMC Scholia
- Alternative chromatin structures of the 35S rRNA genes in Saccharomyces cerevisiae provide a molecular basis for the selective recruitment of RNA polymerases I and II. Goetze H, Wittner M, Hamperl S, Hondele M, Merz K, Stoeckl U, et al. Mol Cell Biol. 2010 Apr;30(8):2028–45. PubMed Europe PMC Scholia
- TOR-dependent reduction in the expression level of Rrn3p lowers the activity of the yeast RNA Pol I machinery, but does not account for the strong inhibition of rRNA production. Philippi A, Steinbauer R, Reiter A, Fath S, Leger-Silvestre I, Milkereit P, et al. Nucleic Acids Res. 2010 Sep;38(16):5315–26. PubMed Europe PMC Scholia
- Casein kinase II-mediated phosphorylation of general repressor Maf1 triggers RNA polymerase III activation. Graczyk D, Debski J, Muszyńska G, Bretner M, Lefebvre O, Boguta M. Proc Natl Acad Sci U S A. 2011 Mar 22;108(12):4926–31. PubMed Europe PMC Scholia
- Recovery of RNA polymerase III transcription from the glycerol-repressed state: revisiting the role of protein kinase CK2 in Maf1 phosphoregulation. Moir RD, Lee J, Willis IM. J Biol Chem. 2012 Aug 31;287(36):30833–41. PubMed Europe PMC Scholia
- Crystal structure of the 80S yeast ribosome. Jenner L, Melnikov S, Garreau de Loubresse N, Ben-Shem A, Iskakova M, Urzhumtsev A, et al. Curr Opin Struct Biol. 2012 Dec;22(6):759–67. PubMed Europe PMC Scholia
- Nutrient sensing and signaling in the yeast Saccharomyces cerevisiae. Conrad M, Schothorst J, Kankipati HN, Van Zeebroeck G, Rubio-Texeira M, Thevelein JM. FEMS Microbiol Rev. 2014 Mar;38(2):254–99. PubMed Europe PMC Scholia