NAD metabolism in oncogene-induced senescence and mitochondrial dysfunction-associated senescence (WP5046)
Homo sapiens

{kind=link}
{kind=link}
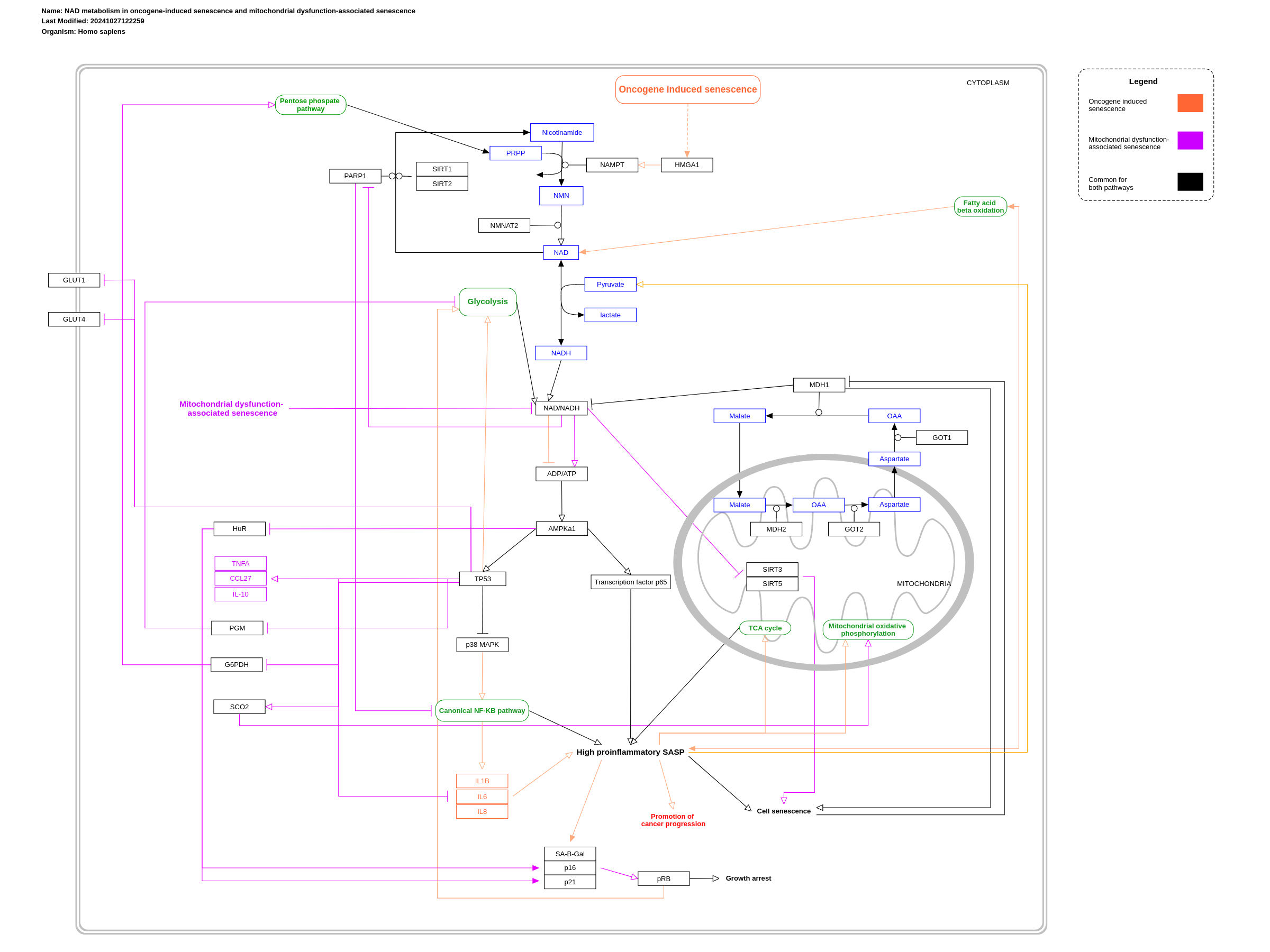
The uppermost part of the pathway includes part of the general NAM salvage pathway in the cytosol as it is relevant to senescence-induced changes to NAD metabolism. In this pathway, NAD levels are maintained through recycling back to NAD from nicotinamide (NAM) and nicotinamide mononucleotide (NMN) (Braidy et al., 2019). The conversion from NAM to NMN is catalyzed by nicotinamide phosphoribosyltransferase (NAMPT), while the conversion from NMN to NAD is catalyzed by nicotinamide mononucleotide adenylyl transferases (NMNATs). Other sources, such as nicotinic acid (NA) and nicotinamid riboside (NR), are not shown here as they are not affected by senescence, at least from current research. OIS-specific interactions are highlighted in orange, while MiDAS-specific interactions are highlighted in purple. General interactions for both (or other senescent types) remain a black color. The OIS pathway, induced by Ras singalling in this case, results in the upregulation of HMGA1, and stimulation of the NAMPT enzyme (Nacarelli et al., 2019). Resulting increased levels of NMN (the direct metabolite of NAMPT) translate to increased NAD levels, and a high NAD-NADH ratio. This leads to decreased ADP-ATP levels, which causes a decreased phosphorylated AMPK expression (Nacarelli et al., 2019). This interaction causes increased p38 and p65 activation, and increased NF-κB activity. The NF-κB signalling pathway has been known to play a key role in the promotion of the proinflammatory SASP (Freund et al., 2011). Furthermore, this is correlated with increased expression of interleukins IL1B, IL6 and IL8, all key factors in the proinflammatory wave of the SASP. In addition, Nacarelli et al. (2019) found that the proinflammatory environment created as a result of the increased NAD-NADH ratio leads to acceleration of cancer progression. NAMPT upregulation through HMGA1 also resulted in the expression of senescence markers SA-ß-gal, p16 and p21. The resulting phenotype from this high NAD-NADH ratio is a high proinflammatory SASP. Malate is another important metabolite in redox reactions and in many senescence types, including OIS and MiDAS. Of interest to NAD metabolism is the malate-aspartate shuttle, where NADH is transferred from the cytosol to the mitochondrial matrix through malate dehydrogenase 1 (MDH1) (Lee et al., 2012). In senescence, levels of MDH1 decrease. On the other hand, decreased activity of MDH1 can induce a senescence response. This reduction in MDH1 activity results in a decreased cytosolic NAD-NADH. Lastly, this inhibition may result in loss of cell proliferation due to the requirement of aspartate synthesis in response to inhibition of the electron transport chain (Birsoy et al., 2015). Mitochondrial dysfunction-associated senescence (MiDAS), on the other hand, causes a decrease in the NAD-NADH ratio, which induces three main responses: (1) the inhibition of sirtuins, (2) the activation of AMPK and (3) the inhibition of PARP which blocks the NF-kB pathway. First, low levels of NAD+ decrease sirtuin activity. A decrease in the activity of SIRT3 and SIRT5, located in the mitochondria, is associated with the activation of cell senescence (Wiley et al., 2016)). Second, a decreased NAD+/NADH ratio activates AMPK and p53, which inhibits the RNA binding protein Hu antigen R (HuR) from degrading the mRNAs encoding the cyclin-dependent kinase inhibitors, p21 and p16INK4a. This increases the activity of the pRB tumor suppressor, resulting in cell proliferation and growth arrest ((Wiley et al., 2016)). Additionally, p53 activation leads to the release of SASPs that lack IL-1-dependent factors but include the secretion of anti-inflammatory cytokine IL-10 and high levels of the pro-inflammatory cytokines CCL27 and TNF-α (Wiley et al., 2016). The activation of p53 also reduces glycolysis and promotes mitochondrial respiration, by inhibiting phosphoglycercate mutase (PGM) and inducing the expression of synthesis of cytochrome c oxidase 2 (SCO2). Furthermore, p53 activation inhibits the pentose phosphate pathway (PPP) by binding to glucose-6-phosphate dehydrogenase (G6PDH). Lastly, the low NAD+,NADH ratio inhibits ADP-ribose donor for poly-ADP ribose polymerase (PARP), which consecutively inhibits the NF-kB pathway. A downregulated NF-κB pathway then contributes to the pathogenic processes of various inflammatory diseases as well as the expression of various proinflammatory SASPs (Liu et al., 2017). As visible in this pathway, when senescence is induced by either OIS or MiDAS distinguishable effects on NAD metabolism are evident. Not only do these stimuli release distinct SASPs, but they exhibit distinct responses on the NAD-NADH ratio and subsequent related pathways. While MiDAS leads to a decrease in the NAD-NADH ratio, OIS causes an increase in this ratio and the NAD+ levels.
For a description of pathway objects, see the WikiPathways Legend.
Authors
Paula Parra , Egon Willighagen , Martina Summer-Kutmon , Kristina Hanspers , Eric Weitz , and Friederike EhrhartActivity

Discuss this pathway
Check for ongoing discussions or start your own.
Cited In
Are you planning to include this pathway in your next publication? See How to Cite and add a link here to your paper once it's online.
Organisms
Homo sapiensCommunities
Annotations
Pathway Ontology
cellular senescence pathway nicotinamide adenine dinucleotide metabolic pathway| Label | Type | Compact URI | Comment |
|---|---|---|---|
| NMN | Metabolite | chebi:16171 | |
| Aspartate | Metabolite | chebi:35391 | |
| Malate | Metabolite | chebi:30797 | |
| OAA | Metabolite | chebi:132560 | |
| PRPP | Metabolite | chebi:17111 | |
| Pyruvate | Metabolite | chebi:15361 | |
| lactate | Metabolite | chebi:24996 | |
| PGM | Metabolite | chebi:33365 | |
| NADH | Metabolite | hmdb:HMDB01487 | |
| NAD | Metabolite | hmdb:HMDB00902 | |
| Nicotinamide | Metabolite | chebi:17154 | |
| IL8 | Metabolite | chebi:138181 | |
| SCO2 | GeneProduct | ensembl:ENSG00000130489 | |
| MDH2 | GeneProduct | ensembl:ENSG00000146701 | |
| GOT2 | GeneProduct | ensembl:ENSG00000125166 | |
| IL1B | GeneProduct | ensembl:ENSG00000125538 | |
| p38 MAPK | GeneProduct | uniprot:Q3C2E3 | |
| HMGA1 | GeneProduct | ensembl:ENSG00000137309 | |
| CCL27 | GeneProduct | ensembl:ENSG00000213927 | |
| GOT1 | GeneProduct | ensembl:ENSG00000120053 | |
| SIRT5 | GeneProduct | ensembl:ENSG00000124523 | |
| IL6 | GeneProduct | ensembl:ENSG00000136244 | |
| NMNAT2 | GeneProduct | ensembl:ENSG00000157064 | |
| PARP1 | GeneProduct | ensembl:ENSG00000143799 | |
| SIRT1 | GeneProduct | ensembl:ENSG00000096717 | |
| SIRT2 | GeneProduct | ensembl:ENSG00000068903 | |
| NAMPT | GeneProduct | ensembl:ENSG00000105835 | |
| TP53 | GeneProduct | ensembl:ENSG00000141510 | |
| SIRT3 | GeneProduct | ensembl:ENSG00000142082 | |
| MDH1 | GeneProduct | ensembl:ENSG00000014641 | |
| TNFA | GeneProduct | ensembl:ENSG000002328 | |
| G6PDH | Protein | ensembl:ENSG00000160211 | |
| GLUT4 | Protein | ensembl:ENSG00000181856 | |
| GLUT1 | Protein | ensembl:ENSG00000117394 | |
| HuR | Protein | ensembl:ENSG00000066044 | |
| Transcription factor p65 | Protein | uniprot:Q04206 | |
| IL-10 | Protein | ensembl:ENSGALG00000000892 | |
| AMPKa1 | Protein | ensembl:ENSG00000132356 |
References
- CD38 and CD157 ectoenzymes mark cell subsets in the human corneal limbus. Horenstein AL, Sizzano F, Lusso R, Besso FG, Ferrero E, Deaglio S, et al. Mol Med. 2009;15(3–4):76–84. PubMed Europe PMC Scholia
- Role of nicotinamide in DNA damage, mutagenesis, and DNA repair. Surjana D, Halliday GM, Damian DL. J Nucleic Acids. 2010 Jul 25;2010:157591. PubMed Europe PMC Scholia
- Senescent cells develop a PARP-1 and nuclear factor-{kappa}B-associated secretome (PNAS). Ohanna M, Giuliano S, Bonet C, Imbert V, Hofman V, Zangari J, et al. Genes Dev. 2011 Jun 15;25(12):1245–61. PubMed Europe PMC Scholia
- Cytosolic malate dehydrogenase regulates senescence in human fibroblasts. Lee SM, Dho SH, Ju SK, Maeng JS, Kim JY, Kwon KS. Biogerontology. 2012 Oct;13(5):525–36. PubMed Europe PMC Scholia
- Retinoblastoma protein promotes oxidative phosphorylation through upregulation of glycolytic genes in oncogene-induced senescent cells. Takebayashi SI, Tanaka H, Hino S, Nakatsu Y, Igata T, Sakamoto A, et al. Aging Cell. 2015 Aug;14(4):689–97. PubMed Europe PMC Scholia
- Mitochondrial Dysfunction Induces Senescence with a Distinct Secretory Phenotype. Wiley CD, Velarde MC, Lecot P, Liu S, Sarnoski EA, Freund A, et al. Cell Metab. 2016 Feb 9;23(2):303–14. PubMed Europe PMC Scholia
- AMPK activation protects cells from oxidative stress-induced senescence via autophagic flux restoration and intracellular NAD(+) elevation. Han X, Tai H, Wang X, Wang Z, Zhou J, Wei X, et al. Aging Cell. 2016 Jun;15(3):416–27. PubMed Europe PMC Scholia
- From Ancient Pathways to Aging Cells-Connecting Metabolism and Cellular Senescence. Wiley CD, Campisi J. Cell Metab. 2016 Jun 14;23(6):1013–21. PubMed Europe PMC Scholia
- NAD+ metabolism governs the proinflammatory senescence-associated secretome. Nacarelli T, Lau L, Fukumoto T, Zundell J, Fatkhutdinov N, Wu S, et al. Nat Cell Biol. 2019 Mar;21(3):397–407. PubMed Europe PMC Scholia